La actividad genética en los micronúcleos: ¿se puede considerar realmente perdido el DNA micronuclear?
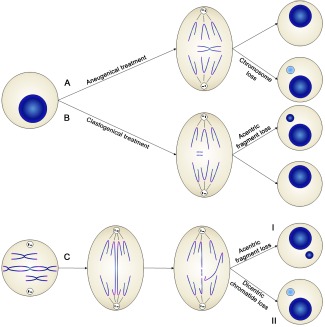
Los micronúcleos son unas estructuras derivadas del núcleo de las células pero, como su nombre indica, son más pequeños que éste. Esto se debe a su origen. En este sentido, el primer mecanismo descrito sobre la formación de micronúcleos se desprendió de un experimento realizado en 1977 por Heddle y Carrano. Estos investigadores observaron que después de la exposición de las células a radiaciones ionizantes, y tras un ciclo de división, las células presentaban micronúcleos. Teniendo en cuenta que la radiación ionizante genera roturas en el DNA, se pudo determinar que los micronúcleos originados contenían fragmentos cromosómicos de tipo acéntrico derivados de roturas no reparadas (Figura1B). Unos años más tarde, Ford y colaboradores observaron que los micronúcleos contenían cromosomas enteros derivados de una mala segregación durante la mitosis (Figura1). Finalmente, los micronúcleos también pueden contener dobles minutos (DM). Los DM son pequeños fragmentos cromosómicos circulares a los que les falta un centrómero activo y los telómeros. Pero su característica principal es que contienen genes altamente amplificados, que a menudo están relacionados con fenotipos cancerígenos (Figura 1C). Años más tarde, Gisselsson y colaboradores, sugirieron que la presencia de micronúcleos y otras anomalías nucleares de tipo morfológico están estrechamente ligadas a las aberraciones mitóticas generadas como consecuencia de los ciclos de rotura, fusión y puente (BFB, breakage-fusion-bridge). Estos ciclos se describieron por primera vez en 1941 (McClintock) y empiezan cuando, ante la presencia de roturas de la cadena del DNA y/o del acortamiento de los extremos cromosómicos (telómeros), los mecanismos de reparación del DNA fallan y se producen fusiones incorrectas entre cromosomas. Estas fusiones favorecen la aparición, durante la división celular, de unas estructuras llamadas puentes anafásicos. Estos puentes se pueden romper y generar otras roturas que, de nuevo, pueden desembocar en la formación de puentes. De esta manera la célula entra en un ciclo persistente de BFB, el que actualmente está considerado como una de las causas principales de inestabilidad cromosómica en cáncer. Por otra parte, al romperse, los puentes también pueden dar lugar a fragmentos acéntrico y a menudo, debido a la tensión generada durante la migración de los cromosomas, estos puentes se pueden despegar. En ambos casos, se pueden acabar generando micronúcleos (Figura1D). Ante estas evidencias, no es de extrañar que los micronúcleos se utilicen de manera rutinaria para detectar la inestabilidad cromosómica.
Mientras que el mecanismo de formación de los micronúcleos está bastante bien definido, el destino de estas estructuras es un tema bastante controvertido. Según Leach y Jackson (2004), existen tres posibles destinos. Uno sería la expulsión del micronúcleo de la célula, lo que llevaría a la pérdida del material genético contenido en el micronúcleo. Otro podría ser la reincorporación de la cromatina del micronúcleo en el núcleo principal de la célula. La última posibilidad, y de hecho la más observada, es la retención del micronúcleo en la célula hasta la siguiente división celular. En este sentido, y suponiendo que el material genético del micronúcleo pudiera segregarse correctamente a uno o ambos núcleos (siempre que su ADN se replicara) de las dos células hijas al final de la división celular, es muy importante que durante la interfase, este material se mantenga intacto. Este hecho, nos llevó a recoger toda la información existente sobre las actividades genéticas en los micronúcleos, es decir, replicación, transcripción y reparación del DNA micronúcleo. Aunque esta información es escasa y a veces contradictoria, intentamos analizarla y encontrar una explicación conforme a todo el mosaico de resultados existentes.
En relación a la replicación del DNA micronúcleo, se encontraron muy pocos estudios que trataran este tema. Sin embargo, se pudo concluir que a pesar de ser poco frecuente, cuando tiene lugar, la replicación del DNA micronuclear se produce en el mismo momento en que tiene lugar la del DNA nuclear, es decir, durante la fase S del ciclo celular (Figura 2).
Figura 2 .- Célula micronucleada con lesiones en el núcleo y los micronúcleos (determinado por la presencia de γH2AX). Mientras que las lesiones de los núcleos presentan el factor de reparación 53BP1, las del micronúcleo no.
En cuanto a la transcripción del DNA micronuclear, los resultados existentes variaban mucho en función del tipo de micronúcleo estudiado en cada caso. Mientras que los micronúcleos que contienen cromosomas enteros presentan una elevada actividad transcripcional, los que contienen fragmentos acéntricos casi no presentan, excepto aquellos que contienen DMs. Estos resultados podrían estar muy ligados a la estructura de la envoltura nuclear del micronúcleo, ya que se ha observado que éste es diferente según el tipo de micronúcleo. En el núcleo principal, la envoltura nuclear está formado por dos membranas lipídicas que se unen entre ellas en unos puntos llamados poros nucleares. Estos poros permiten el transporte de moléculas entre el núcleo y el citoplasma. En cuanto a los micronúcleos, mientras que aquellos que contienen cromosomas enteros presentan una envoltura muy parecida al del núcleo principal, los que contienen fragmentos acéntricos presentan una envoltura incumplido. Una posible explicación podría radicar en el momento de la formación de este envoltorio. Algunos autores han especulado que la formación de poros nucleares depende de la unión de algunas de sus proteínas estructurales con secuencias específicas del DNA (Sukegawa y Blobel). Así pues, el hecho de que un micronúcleo contenga estas secuencias, favorecería la correcta formación de su envoltorio. Teniendo en cuenta que los micronúcleos con cromosomas enteros contienen más material genético que aquellos que sólo contienen fragmentos cromosómicos, las posibilidades que dentro del micronúcleo se puedan encontrar estas secuencias son más elevadas y, en consecuencia, la probabilidad de que el envoltorio sea completo también . Sin embargo, este razonamiento no permitiría explicar por qué los micronúcleos que contienen DMs pueden transcribir su DNA con más facilidad que los que contienen fragmentos acéntricos no circulares. En este sentido, cabe destacar que los micronúcleos de tipo DM están sujetos a una alta presión selectiva ya que la presencia de las amplificaciones génicas contenidas en estos micronúcleos son decisivas para la supervivencia de las células que las contienen. Por lo tanto, harían falta más estudios para poder explicar por qué los micronúcleos de tipo DM se comportan de manera diferente a los micronúcleos que contienen fragmentos acéntricos convencionales.
Finalmente, muchas publicaciones presentan resultados haciendo referencia a la presencia de factores reparación del DNA en micronúcleos radio-inducidos. Como ya se ha dicho, este tipo de micronúcleos se generan básicamente debido a la no reparación de las dobles roturas de cadena. Por este motivo, estos estudios se centran básicamente en el estudio de los factores de reparación de las vías que reparan las dobles roturas de cadena (Haaf, Medvedeva, Yoshikawa). Los factores de estudio varían en función del trabajo, pero la observación más relevante es la ausencia de proteínas de reparación en lugares donde hay roturas. Por otra parte y, de manera no menos importante, mientras que algunos estudios claman la presencia de ciertos factores, otros resaltan su ausencia. Teniendo en cuenta la experiencia de nuestro grupo en este tema (Terradas y colaboradores, 2009), creemos que la causa de esta diversidad de resultados podría ser un defecto en el transporte de los factores de reparación hacia el micronúcleo. En este sentido, ya hemos comentado que la envoltura nuclear de los micronúcleos con fragmentos acéntricos no era del todo completa (Hoffelder). Además, los trabajos realizados en nuestro laboratorio, nos permitieron observar que una parte de estos micronúcleos presentan marcadores de degradación del DNA. El aumento de la fracción de micronúcleos en vías de degradación se producía en detrimento de la disminución de los micronúcleos que presentaban marcadores de daño. Por este motivo, creemos que aquellos micronúcleos donde su DNA no se ha podido reparar podrían acabar degradándose.
En conclusión, la naturaleza de los micronúcleos está altamente relacionada con su destino final. En aquellos micronúcleos donde la estructura de su envoltura sea idéntica a la del núcleo principal, las actividades genéticas aquí revisadas se podrán producir. De este modo, se favorecerá el mantenimiento de estas estructuras hasta la siguiente división celular. En cambio, aquellos micronúcleos con una envoltura incumplido serán más sensibles a la pérdida de integridad durante la vida celular, incrementando aún más la inestabilidad genómica de las células micronucleades, en este caso, en términos de dosis génica.
Referencias
"Genetic activities in micronuclei: Is the DNA entrapped in micronuclei lost for the cell?". Terradas, Mariona; Martin, Marta; Tusell, Laura; Genesca, Anna. MUTATION RESEARCH, 705 (1): 60-67 (2010).